Share this project

Martijn Bart
Universiteit van Amsterdam
Publication date: 30 juni 2021
University: Universiteit van Amsterdam
ISBN: 978-94-91407-97-0
BRINGING LIFE TO THE DEEPEST FRONTIER
Summary
The deep sea forms by far the largest part of our biosphere, is the largest reservoir of biomass on the planet, and is home to the largest number of (undiscovered) species. Yet, the deep sea is generally considered an inhospitable environment because of the cold temperatures, high pressure, and limited light availability under which deep-sea organisms have to survive. The aforementioned conditions also put technical constraints on the possibilities to study deep-sea organisms and their ecosystems. Consequently, the majority of deep-water ecosystems is still relatively undescribed while at the same time the human imprint on these ecosystems is unmistakably increasing via bottom trawling, deep-sea mining, oil and gas exploration and climate change. In recent years, the rapid development of sophisticated technology (e.g., remotely operated vehicles (ROVs) and high resolution imaging) has improved the opportunities to observe and study deep-sea ecosystems. Particularly in the North-Atlantic Ocean, the seafloor is abundantly inhabited by sponges that form large mono-specific sponge grounds, create sponge reefs and are major components of deep-sea coral reefs. These sponge ecosystems increase local biodiversity and productivity, and have been suggested to play a crucial role in the (re)cycling of elements such as carbon, nitrogen and silica. The aim of this thesis is to assess the carbon and nitrogen cycling through dominant deep-sea sponge species, and to evaluate the ecological impact of resource cycling within deep-sea sponge ecosystems.
Sponges, including deep-sea species, acquire food by filter feeding. They actively pump large amounts of water through their bodies and efficiently capture and process food particles, such as bacterioplankton. Additionally, it has been shown that many shallow-water sponges primarily rely on dissolved organic matter (DOM) as a food source. DOM, often measured in the form of dissolved organic carbon (DOC), is the largest potential food source in the oceans, but generally represents only a minor fraction of the food intake of most invertebrate organisms. Yet, shallow water sponges possess the remarkable capacity to almost completely fulfill their nutritional needs by DOM-feeding — usually DOM represents more than 90 % of their daily organic carbon intake. Using ex situ (in a laboratory aquarium set-up) and in situ (at the seafloor) incubations, we found that various dominant deep-sea sponge species also rely heavily on DOM as a food source, irrespective of the sponge’s phylogeny, growth form, its abundance and composition of microbial symbionts, or differences in initial ambient plankton and DOC concentrations. For four different dominant North-Atlantic deep-sea sponge species, DOC was found to represent 92–100 % of their total net organic carbon removal, and is therefore a crucial food source for these sponges to sustain themselves on the otherwise food-limited ocean floor.
Sponges live in association with a variety of microorganisms and are therefore considered ‘holobionts’. Generally, sponges are classified as having either low microbial abundances (LMA sponges) or high microbial abundances (HMA sponges). Microbial symbionts are considered crucial to the functioning of the holobiont as they are involved in various metabolic processes, such as carbon and nitrogen metabolism. To study how various sponge types process (i.e. assimilate and respire) their food, we used 13C- and 15N-enriched DOM and bacterioplankton as food sources for these sponges in ex situ incubation experiments. We showed that whilst LMA sponges quantitatively take up more DOC, a relatively high amount of this DOC is lost as CO2 during respiration. In contrast, HMA sponges assimilate a higher percentage of DOC into sponge biomass, rather than lose it through respiration, and can thus be considered more efficient in processing DOM. This indicates that functional traits (e.g., abundance of microbial symbionts and morphology) influence the food preferences and diet composition of sponges, and that deep-sea sponges use various ‘survival strategies’ to thrive on the seafloor. Furthermore, in all of the investigated sponge species assimilation-to-respiration efficiencies were higher for the bacterioplankton food source than for DOM. Thus, although DOM constitutes a much larger proportion of the daily diet of sponges compared to bacterioplankton, the latter food source seems to be preferred. Based on these results we hypothesize that both food sources serve different purposes for sponge nutrition: DOM serves as the main energy source for deep-sea sponges to sustain their minimal energetic requirements, while supplementation with bacteria and other high-quality particulate food sources is essential to support anabolic processes (e.g., somatic growth, reproduction, and cell turnover).
We then integrated our organic and inorganic carbon and nitrogen measurements into an organismal linear inverse network model for the species Geodia barretti, covering a range of individual sponge sizes (10‒3500 mL), to quantify both known and unknown carbon and nitrogen metabolic processes, including respiratory quotients, production efficiencies, carbon fixation and production rates (P/B), and nitrogen loss as N2. The model revealed that the DOM taken up by G. barretti is primarily used for aerobic respiration, then for dissimilatory nitrate (NO3-) reduction to ammonium (NH4+) (DNRA), and last, for denitrification. Internally produced NH4+ from cellular excretion and DNRA fueled nitrification and contributed up to 7 % of total sponge biomass production. Furthermore, the overall relative pattern of carbon and nitrogen metabolism proved rather independent of sponge size (i.e. the dominant processes were similar between larger and smaller sized individuals), but metabolic rates were higher in smaller sponges, indicating they use more energy for growth than larger specimens. Using our model estimated RQ and C:N conversions for sponges of different size classes, future studies can now approximate C and N fluxes based on in situ assessed O2 fluxes alone. Metabolic modelling of hard-to-reach, deep-water sponges can be used to predict community-based biogeochemical fluxes and sponge production, especially when only a subset of in situ parameters are known, and can facilitate further investigations on the ecological significance of the sponge aggregations in deep-sea ecosystems.
On shallow tropical coral reefs, the capacity of sponges to take up DOM does not only provide them with a valuable addition to their diet, but also makes them important players in the recycling of energy. A large fraction of the DOM that shallow-water sponges take up is invested in a constant renewal of their filter cells, whilst simultaneously, damaged or old filter cells are released from the sponge as ‘sponge poo’ or detritus. This sponge poo was subsequently discovered to be an important food source for various detritivorous organisms living on the reef. This process of DOM-feeding by sponges, followed by detritus production and detritus feeding by other organisms, was termed the ‘sponge loop’. Carbon fluxes through the sponge loop were estimated to be in the same order of magnitude as primary production rates of the entire coral reef ecosystem and therefore provided an important key to resolve Darwin’s 180-year old paradox of how coral reefs act as oases of biodiversity and productivity in otherwise desert-like waters. However, while efficient recycling of energy could also be crucial in food limited deep-sea environments, the sponge-loop pathway has never been verified in deep-sea ecosystems. To investigate the release and transfer of processed dissolved and particulate organic carbon and nitrogen from deep-sea sponges to associated deep-sea fauna, we performed a pulse-chase experiment using 13C- and 15N-enriched food sources. We found that particulate detritus released by two sponge species contained tracer-13C from the previously consumed tracer DOM and POM, and, after 9 days of exposure to the labeled sponges and detritus, enrichment of 13C and 15N was also detected in the tissue of associated brittle stars. These results therefore provide the first evidence of a complete sponge-loop pathway in the deep sea. However, to assess the ecological importance of sponge-loop pathways in the deep sea — both qualitatively and quantitatively — many questions remain to be answered. It is for example still unclear how metabolic rates of deep-sea sponges affect cell turnover and detritus production, via which mechanisms various sponges with different functional traits (e.g., abundance of microbes, growth form) transfer organic material to higher trophic levels (i.e. via the detritivorous or predatory pathway), and what the quantitative importance is of sponge-loop pathways in the wide variety of deep-sea ecosystems in which sponges thrive.
The future challenges of deep-sea (sponge) biology are not restricted to science alone. Present-day deep-sea research experiences a strong connection with the commercial exploration of natural resources. A focus on resources that provide prospective returns on investments can result in a neglect of other research opportunities that are not directly useful towards that investment. Consequently, while we commercially exploit the deep-sea for fish, minerals, or oil and gas, the basic scientific knowledge required to understand deep-sea ecosystems in order to preserve or exploit them is lacking. To increase our knowledge baseline of the deep sea and to understand the impact of anthropogenic stressors on deep-sea ecosystems, independent scientific investigations are essential. Preferably, these studies are presented in ways that appeal to the general public — for example by touching broader interdisciplinary topics such as the origin of life — in order to change the perception of the deep sea as a monstrous inhospitable place to that of a vital and ethically-relevant ecosystem. This is needed to turn scientific results into active conservation- and protection measures, because we as humans are more inclined to protect something we feel connected to and care about. As a relatively new scientific discipline, deep-sea biology can use examples of other scientific fields related to conservation (e.g., ‘the land ethic’ and the red-listing of species) and outreach (e.g., ‘Earthrise’ and the science of space exploration) to its advantage. Moreover, a better integration of deep-sea biology with disciplines such as philosophy and ethics can help to put experimental results in perspective and stimulate environmental awareness, while active collaborations with law- and policy makers are needed to provide a legal framework for deep-sea exploitation and to put potential conservation measures into practice.
See also these dissertations


Toby van Gastelen
Structure-Preserving Data-Driven Methods for Modeling Turbulent Flows
19 mei 2026
TU Eindhoven
Open Ebook
Ton Winkelmolen
Molecular insights into the role of VRS5 in tillering and lateral spikelet development in barley
12 juni 2026
Wageningen University
Open Ebook
Selcuk Peker
Gamma Knife Radiosurgery for Skull Base Tumors
10 juni 2026
Universiteit Maastricht
Open Ebook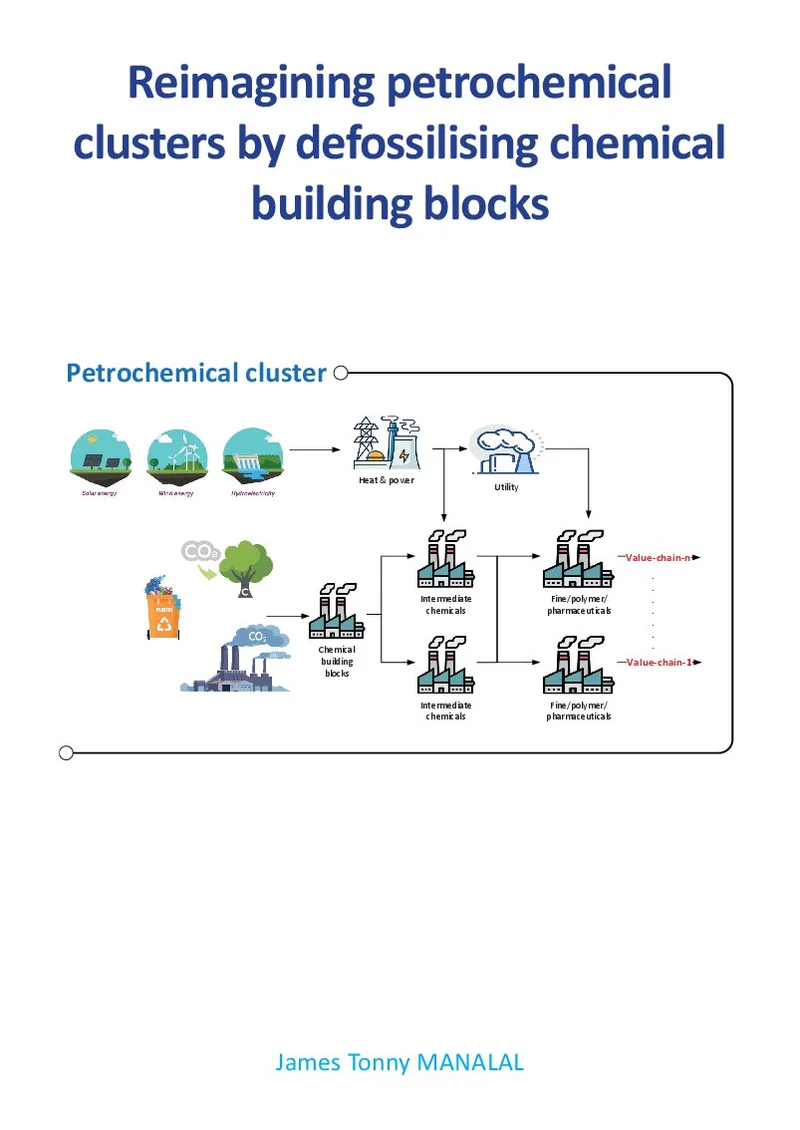
Tonny Manalal
Reimagining petrochemical clusters by defossilising chemical building blocks
18 mei 2026
Overig
Open Ebook



Cora De Gol
Microbial stabilization and protein functionality of plant-based liquids using pulsed electric fields
17 juni 2026
Wageningen University
Open Ebook
We print for the following universities


